-
染色体
 染色体
染色体
染色体(chromosome)是细胞在有丝分裂或减数分裂时DNA存在的特定形式,由染色质丝螺旋缠绕,逐渐缩短变粗形成。
染色体有种属特异性,随生物种类、细胞类型及发育阶段不同,其数量、大小和形态存在差异。
目录
- 1 基本信息
2 基本简介
3 发现时期
4 结构简介
5 基本组型
6 结构序列
7 不同状态
8 不同生物
9 性染色体
10 全面分析
11 异常情况
12 最新进展
13 变异情况
14 细胞起源
基本信息
外文名:Chromosome
意义:遗传信息的载体
中文名:染色体
组成:主要为DNA、蛋白质
分布:真核细胞
基本简介
 染色体
染色体
在无性繁殖物种中,生物体内所有细胞的染色体数目都一样;而在有性繁殖大部分物种中,生物体的体细胞染色体成对分布,含有两个染色体组,称为二倍体。
性细胞如精子、卵子等是单倍体,染色体数目只是体细胞的一半。哺乳动物雄性个体细胞的性染色体对为XY,雌性则为XX。鸟类.两栖类.爬行类和某些昆虫的性染色体与哺乳动物不同:雄性个体的是ZZ,雌性个体为ZW。
发现时期
1879年德国生物学家弗莱明(Fleming·w)把细胞核中的丝状和粒状的物质,用染料染红,观察发现这些物质平时散漫地分布在细胞核中,当细胞分裂时,散漫的染色物体便浓缩,形成一定数目和一定形状的条状物,到分裂完成时,条状物又疏松为散漫状。
1888年正式被命名为染色体。
 染色体
染色体
1928年摩尔根通过果蝇杂交实验证实了染色体是基因的载体,从而获得了生理医学诺贝尔奖。
1953年4月《自然》杂志刊登了美国的沃森和英国的克里克在英国剑桥大学合作的研究成果:DNA双螺旋结构的分子模型,被誉为20世纪以来生物学方面最伟大的发现。
1956年,美籍华裔遗传学家Joe Hin Tjio(1919–2001,资料译为庄有兴或蒋有兴)和Levan首次发现人的体细胞的染色体数目为46条,标志着人类细胞遗传学的建立。46条染色体按其大小、形态配成23对,第一对到第二十二对叫做常染色体,为男女共有,第二十三对是一对性染色体(Sex Chromosome),雄性个体细胞的性染色体对为XY;雌性则为XX。
结构简介
染色体的超微结构显示染色体是由直径仅100埃(Å,1埃=0.1纳米)的DNA-组蛋白高度螺旋化的纤维所组成。每一条染色单体可看作一条双螺旋的DNA分子。有丝分裂间期时,DNA解螺旋而形成无限伸展的细丝,此时不易为染料所着色,光镜下呈无定形物质,称之为染色质。有丝分裂时DNA高度螺旋化而呈现特定的形态,此时易被碱性染料着色,称之为常染色体。
1970年后陆续问世的各种显带技术对染色体的识别作出了很大贡献。中期染色体经过DNA变性、胰酶消化或荧光染色等处理,可出现沿纵轴排列的明暗相间的带纹。按照染色体上特征性的标志可将每一个臂从内到外分为若干区,每个区又可分为若干条带,每条带又再分为若干个亚带,例如“9q34.1”即表示9号染色体长臂第3区第4条带的第1个亚带。由于每条染色体带纹的数目和宽度是相对恒定的,根据带型的不同可识别每条染色体及其片段。
80年代以来根据DNA双链互补的原理,应用已知序列的DNA探针进行荧光原位杂交(Fluorescence in situ hybridization,FISH)可以识别整条染色体、染色体的1个臂、1条带甚至一个基因,因而大大提高了染色体识别的准确性和敏感性。染色体是遗传物质—基因的载体,控制人类形态、生理和生化等特征的结构基因呈直线排列在染色体上。2000年6月26日人类基因组计划(HGP)已宣布完成人类基因组序列框架图。2001年2月12日HGP和塞雷拉公司公布了人类基因组图谱和初步分析结果。人类基因组共有3~3.5万个基因,而不是以往认为的10万个。由此可见,染色体和基因二者密切相关,染色体的任何改变必然导致基因的异常。
成分
染色体的主要化学成份是脱氧核糖核酸(DNA)和蛋白质构成,染色体上的蛋白质有两类:一类是低分子量的碱性蛋白质即组蛋白(histones),另一类是酸性蛋白质,即非组蛋白蛋白质(non-histone proteins)。非组蛋白蛋白质的种类和含量不十分恒定,而组蛋白的种类和含量都很恒定,其含量大致与DNA相等。所以人们早就猜测,组蛋白在DNA·蛋白质纤丝的形成上起着重要作用。Kornberg根据生化资料,特别是根据电镜照相,最先在1974年提出绳珠模型(beads on-a-string model),用来说明DNA·蛋白质纤丝的结构。纤丝的结构单位是核小体,它是染色体结构的最基本单位。
核小体的核心是由4种组蛋白(H2A、H2B、H3和H4)各两个分子构成的扁球状8聚体。我们知道,DNA分子具有典型的双螺旋结,一个DNA分子就像是一条长长的双螺旋的纤丝。一条染色体有一个DNA分子。DNA双螺旋依次在每个组蛋白8聚体分子的表面盘绕约1.75圈,其长度相当于140个碱基对。组蛋白8聚体与其表面上盘绕的DNA分子共同构成核小体。在相邻的两个核小体之间,有长约50~60个碱基对的DNA连接线。在相邻的连接线之间结合着一个第5种组蛋白(H1)的分子。密集成串的核小体形成了核质中的100埃左右的纤维,这就是染色体的“一级结构”,就像成串的珠子一样,DNA为绳,组蛋白为珠,被称作染色体的“绳珠模型”如图→在这里,DNA分子大约被压缩了7倍。
 染色体
染色体
300埃左右的螺线体(二级结构)再进一步螺旋化,形成直径为0.4微米(μm)的筒状体,称为超螺旋管。这就是染色体的“三级结构”。到这里,DNA又再被压缩了40倍。超螺旋体进一步折叠盘绕后,形成染色单体—染色体的“四级结构”。两条染色单体组成一条染色体。到这里,DNA的长度又再被压缩了5倍。从染色体的一级结构到四级结构,DNA分子一共被压缩了7×6×40×5=8400倍。例如,人的染色体中DNA分子伸展开来的长度平均约为几个厘米,而染色体被压缩到只有几纳米长。
基本组型
染色体组型(Karyotype):描述一个生物体内所有染色体的大 小、形状和数量信息的图象。这种组型技术可用来寻找染色体歧变同特定疾病的关系,比如:染色体数目的异常增加、形状发生异常变化等。以染色体的数目和形态来表示染色体组的特性,称为染色体组型。虽然染色体组型一般是以处于体细胞有丝分裂中期的染色体的数目和形态来表示,但是,也可以其他时期,特别是以前期或分裂间期的染色体形态来表示。
小、形状和数量信息的图象。这种组型技术可用来寻找染色体歧变同特定疾病的关系,比如:染色体数目的异常增加、形状发生异常变化等。以染色体的数目和形态来表示染色体组的特性,称为染色体组型。虽然染色体组型一般是以处于体细胞有丝分裂中期的染色体的数目和形态来表示,但是,也可以其他时期,特别是以前期或分裂间期的染色体形态来表示。
关于整个染色体的情况可作下列记载而加以表示:各自的长度、粗细;着丝粒的位置;随体及次缢痕的有无、数目、位置;凝缩部不同的部分以及异染色质部分、常染色质部分;染色粒、端粒的形态、大小及分布情况;小缢痕的数目、位置;由于温度和药品处理所产生的染色体分带(band)的形态、数目、位置等等。
对于染色体组的表示,现已提出几种方法。例如,染色体的数目是以n、2n分别表示配子和合子的染色体数目,以x表示基数,以b表示原始基数,以2x、3x、4x、……表示多倍性,以2x 1、2x-1、……等等表示非整惰性,以1、2、3、……等编号表示各个染色体。
另外,为了表示各个染色体的形态特征,还可采用“V”形、“J”形等名称,或者采用由A.Levan等(1964)所提出的根据着丝粒的位置进行分类的方法等。关于人类的染色体组型的表示法,在国际上是统一的(在丹佛1960、伦敦1963、芝加哥1966、巴黎1971等地召开的人类染色体会议上所制订的),已规定了为了表示染色体形态特征的染色体臂比、着丝点指数等指标。
结构序列
染色体要确保在细胞世代中保持稳定,必须具有自主复制、保证复制的完整性、遗传物质能够平均分配的能力,与这些能力相关的结构序列是:
自主复制
20世纪70年代末首次在酵母菌中发现。自主复制DNA序列具有一个复制起始点,能确保染色体在细胞周期中能够自我复制,从而保证染色体在世代传递中具有稳定性和连续性。
着丝粒
着丝粒DNA序列与染色体的分离有关。着丝粒DNA序列能确保染色体在细胞分裂时能被平均分配到2个子细胞中去。
着丝粒DNA序列特点:
(1)一方面在所有的真核生物中它们的功能是高度保守的,另一方面即使在亲缘关系非常相近的物种之间它们的序列也是多样的。
(2)绝大多数生物的着丝粒都是由高度重复的串联序列构成的,然而,在着丝粒的核心区域,重复序列的删除,扩增以及突变发生的非常频繁,种种研究表明,重复序列并不是着丝粒活性所必须的。
(3)有些科学家提出了可能是DNA的二级结构甚至是高级结构是决定着丝粒位置和功能的因素。即功能的序列无关性。
端粒
为一段短的正向重复序列,在人类为TTAGGG的高度重复序列。端粒DNA功能是保证染色体的独立性和遗传稳定性。
染色体的分裂分三种; 一是母钟分裂,这个一般发生在受精卵的早期,人类具体就是从一条受精卵分裂为个体的23对染色体的过程,意思是按照母体蓝图进行子代分裂,被分裂的23对染色体分别可以造出各种组织器官,如果第一条是造肝的,那么它上面的所有造肝的基因片段都被打开,相反其它器官的制造信息都被关闭,这个过程母体蓝图染色体要分裂4次(按几何级数分裂);
一是母钟分裂,这个一般发生在受精卵的早期,人类具体就是从一条受精卵分裂为个体的23对染色体的过程,意思是按照母体蓝图进行子代分裂,被分裂的23对染色体分别可以造出各种组织器官,如果第一条是造肝的,那么它上面的所有造肝的基因片段都被打开,相反其它器官的制造信息都被关闭,这个过程母体蓝图染色体要分裂4次(按几何级数分裂);
二是子钟分裂,按照母体蓝图分裂的23对人类染色体已经在“母钟分裂”过程中分别被打开,它们各自按照各自的“子代蓝图”进行下面造器官的分裂,一个个有机的器官从此被造出,并且开始发挥各自的功能,这个过程子体蓝图染色体要分裂24次(个物种染色体的不同,其分裂的次数也不同,不过一个总的原则是按染色体数分裂),在24次分裂后,一个完整的人体就被造出来;三是孙钟分裂,一个独立的人体,在生长发育的过程中,还有一些器质性和功能性的东西没有出现,所以必须再打开,进行再分裂。比如七岁儿童脱牙,十多岁少年具有生育能力,有些遗传病到一定时候的发作,等等。
对应三种分裂,必须有三种控制分裂发生的手段。母钟分裂是“端点(又叫端粒)控制体系”,这种分裂的原始触发点在外界,比如飘荡在空气中的细菌,它只要没有接触食物或易感物,就永远是不产生分裂的原命,一旦接触,在端点的作用下就开始母钟分裂。子钟分裂是受制于子钟染色体的端点,与外界刺激无关。孙钟染色体分裂受制于染色体外相对应的一些蛋白质,它们的功能仅仅是到一定时间将这个包含某信息的片段打开。
依此看来,染色体就是人体的生物钟。所以我们将第一条受精卵叫“母钟”,将母钟分裂出来的23对染色体叫子钟,将23对染色体造出的各种组织器官所包含的染色体叫“孙钟”,改变子钟、孙钟的染色体都不可以改变遗传,只有改变母钟的基因才可以造成“变异”。
染色体可以携带“遗传基因”但是不能传递“打开信息”,打开某个基因段的所有信息都是通过染色体端点或染色体外的蛋白质发挥作用才完成分裂或复制的。分裂是染色体整体的,复制是染色体某个基因片段的。
不同状态
基因在细胞里并非一盘“散沙”或“散兵游勇”,它们大多有规律地集中在细胞核内的染色体上,而且每一种生物细胞内染色体的形态和数目都是一定的。
染色体复制时
染色体在复制以后,含有纵向并列的两个染色单体(c hroma-tids),只有在着丝粒(centromere)区域仍联在一起。着丝粒在染色体上的位置是固定的。由于着丝粒位置的不同,把染色体分成大致相等或长短不等的两臂(arms)。着丝粒的位置在染色体中间或中间附近时,染色体两臂的长度差不多,这着丝粒叫做中间着丝粒或亚中间着丝粒。着丝粒的位置靠近染色体的一端时,根据着丝粒离开端部的远近,这着丝粒叫做近端部着丝粒或端部着丝粒。着丝粒所在的地方往往表现为一个缢痕,所以着丝粒又称初级缢痕(primary constriction)。
hroma-tids),只有在着丝粒(centromere)区域仍联在一起。着丝粒在染色体上的位置是固定的。由于着丝粒位置的不同,把染色体分成大致相等或长短不等的两臂(arms)。着丝粒的位置在染色体中间或中间附近时,染色体两臂的长度差不多,这着丝粒叫做中间着丝粒或亚中间着丝粒。着丝粒的位置靠近染色体的一端时,根据着丝粒离开端部的远近,这着丝粒叫做近端部着丝粒或端部着丝粒。着丝粒所在的地方往往表现为一个缢痕,所以着丝粒又称初级缢痕(primary constriction)。
有些染色体上除了初级缢痕以外,还有一个次级缢痕(secon-dary constriction),连上一个叫做随体(satellite)的远端染色体小段。次级缢痕的位置也是固定的。在细胞分裂将结束时,核内出现一个到几个核仁,核仁总是出现在次级缢痕的地方,所以次级缢痕也叫做核仁形成区(nucleolar organizer)如图:着丝粒是有丝分裂或减数分裂中的染色体高度压缩的一个区域,在此纺锤体纤维与其结合。复杂的DNA序列构成了着丝粒。发育的面包酵母(啤酒酵母)的着丝粒长度约为220个碱基对,并且通过多种与剩余DNA结合组蛋白有明显区别的蛋白质共同保护其免受限制性核酸内切酶(简称限制酶)的消化。虽然着丝粒区域通过特殊的蛋自质保护其免受限制性内切核酸酶的攻击,但该区域投有核小体而且被去凝聚,这似乎说明了在有丝分裂和减数分裂过程中着丝粒区域被高度缩窄原因。着丝粒的220对碱基序列两侧是限制性内切核酸酶敏感位点,该位点的功能也许是促进DNA的断裂,有助于染色单体在后期的相互分离。限制性内切核酸酶是一种在核酸内特殊位点进行切割的酶类。
不同生物
真核生物的基因分布在许多染色体中,一般来讲这些染色体在大小上有很大不同。与细菌染色体(由环状DNA分子构成)比较,真核染色体含有线性双链DNA。DNA和多种类型的相关蛋白质构成r染色体。真核染色体的结构成分中并没有RNA。
真核染色体可被不同程度的浓缩。最低的浓缩结构是伸展的核小体形式(extended nucleosome form)(图3一la)。核小体是由近乎球状的组蛋白形成的八聚体(histone)(H2a、H2b、H3和H4)和在其外围绕两圈的DNA所构成。DNA的约200个碱基对围绕着由组蛋白构成的球形体。并有DNA的50个碱基对连接相邻的核小体。虽然真核染色体伸展的核小体形式与细菌当中看到的串珠样的结构相似,但是这些结构可能并不相同。伸展的核小体形式存在于染色体将被复制的区域,或存在于与RNA分子合成有关的区域。真核DNA更紧密的状态是螺线管形式(solenoid form)。与核小体结合的组蛋白H1诱导其组装成6个核小体的环,并且这些环组成圆筒状螺线管结构。在分裂间期大多数真核染色体以螺线管形式存在。进行复制或被表达的(转录为RNA分子)这些部分去解凝成为伸展的核小体形式。DNA的复制发生在间期的S阶段。基因表达发生在间期的所有阶段(G0、S和G1)。那些不进行增殖的(进入细胞周期)真核细胞被认为是在G0阶段,并且与间期细胞相似。在间期细胞核中看到的染色质是由绝大多数以螺线管形式存在的DNA 构成。
DNA最紧密的状态是环状的螺线管形式。DNA结合蛋白促进螺线管在支架蛋白中心核前后形成环状。在一些真核生物中螺线管的l8个环组成了一个盘状结构。染色体凝聚为数百个叠在一起的盘状结构。在有丝分裂和减数分裂的过程中,可观察到环状的螺线管形式。由于许多长的染色体必须在细胞内移动,并且在移动过程中可能被牵扯,所以染色体的浓缩是必要的。
原核细菌
在高中阶段,染色体必须由DNA和蛋白质组成。由于原核生物的染色体一般不含组蛋白,所以高中阶段称原核生物没有染色体。
一般而言,原核生物的染色体可以进行复制,但大多数细胞容易存活多份。
所有必需的细菌基因存在于细胞质中的单个环状双链DNA(dsDNA)染色体中。细菌染色体与质膜相附着。细菌染色体(bacterial chromosome)依其种类不同可编码1000个或5000 个蛋白质。除了细菌染色体以外,还可有一个或多个较小的染色体,称为质粒,它一般指定 20一100个蛋白质。质粒是环状双链DNA分子,它可与或不与质膜附着。质粒编码的大多数或全部蛋白质在正常环境条件下并不是细胞生存所绝对必需的。许多质粒编码的蛋白质使其把一些遗传信息向其他细胞转移成为可能,并促进稀有化合物的代谢。或使细胞可抵抗某些化学物质或重金属。
由DNA、蛋白质和RNA构成的细菌染色体是高度浓缩的。它不仅通过拓扑异构酶(topoisomerase)形成超螺旋,并且,环绕在由RNA和蛋白质形成的“核”的内外。许多DNA的负电荷被多胺和DNA缠绕着的碱性蛋白质所中和。通过柔和地裂解细菌细胞得到的DNA外观呈串珠状。虽然细菌染色体也是高度浓缩的,但是,在光学显微镜下它们不能被看到。在透射电子显微镜下,细菌染色体的外观与非分裂的真核细胞核内的染色质(chiromatin)非常相像。
人类染色体
男性体细胞染色体的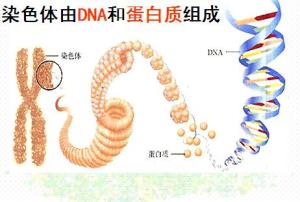 组成:22对常染色体+XY。女性体细胞染色体的组成:22对常染色体+XX。但是,男性生殖细胞染色体的组成:22条常染色体+X或Y。女性生殖细胞染色体的组成:22条常染色体+X。
组成:22对常染色体+XY。女性体细胞染色体的组成:22对常染色体+XX。但是,男性生殖细胞染色体的组成:22条常染色体+X或Y。女性生殖细胞染色体的组成:22条常染色体+X。
人体的体细胞内有23对染色体。包括22对常染色体和一对性染色体。性染色体包括:X染色体和Y染色体。含有一对X染色体的受精卵发育成女性,而具有一条X染色体和一条Y染色体者则发育成男性。这样,对于女性来说,正常的性染色体组成是XX,男性是XY。这就意味着,女性细胞减数分裂产生的配子都含有一个X染色体;男性产生的精子中有一半含有X染色体,而另一半含有Y染色体。精子和卵子的染色体上携带着遗传基因,上面记录着父母传给子女的遗传信息。同样,当染色体异常时,就可形成遗传性疾病。男性不育症中因染色体异常引起者约占2%~21%,尤其以少精子症和无精子症多见。
数目列表
每一种生物的染色体数是恒定的。多数高等动植物是二倍体(diploid),也就是说,每一身体细胞中有两组同样的染色体(有时与性别直接有关的染色体,即性染色体,可以不成对)。亲本的每一配子带有一组染色体,叫做单倍体(haploid),用n来表示。两个配子结合后,具有两组染色体,叫做二倍体,用2n表示。例如玉米的二倍体染色体数是20(2n=20),即有10对染色体,如图→。人的染色体数是46(2n=46),即有23对染色体,如图←。但多数微生物的营养体是单倍体,例如链孢霉的单倍体染色体数是7 。
| 通名 | 学名 | 双倍体数 |
|---|---|---|
| 动物 | ||
| 人类 | Homo sapiens | 46 |
| 弥猴 | Macacamalatta | 42 |
| 黄牛 | Bostaurus | 60 |
| 猪 | Susscrofa | 38 |
| 狗 | Canis familiaris | 78 |
| 猫 | Felis domesticus | 38 |
| 马 | Equus Calibus | 64 |
| 驴 | Equus asinus | 62 |
| 山羊 | Capara hircus | 60 |
| 绵羊 | Ovis aries | 54 |
| 小家鼠 | Mus musculus | 40 |
| 大家鼠 | Rattus norvegicus | 42 |
| 水貂 | Mustela vison | 30 |
| 豚鼠 | Cavia cobaya | 64 |
| 兔 | Oryctolagus cuniculus | 44 |
| 家鸽 | pigeon | 约80 |
| 鸡 | Gallus domesticus | 约78 |
| 火鸡 | Meleagris gallopavo | 约80 |
| 鸭 | Anas platyrhyncho | 约80 |
| 家蚕 | Bombyx mori | 56 |
| 家蝇 | Musca domestica | 12 |
| 果蝇 | Drosophila melanogaster | 8 |
| 蜜蜂 | Apis mellifera | ♀32♂16 |
| 蚊 | Culex pipiens | 6 |
| 佛蝗 | Phlaeobainfumata | ♀24♂23 |
| 水螅 | hydra | 32 |
| 植物 | ||
| 洋葱 | Alliumcepa | 16 |
| 大麦 | Hordeum uulgare | 14 |
| 水稻 | Oryza sativa | 24 |
| 小麦 | Triticum vulgare | 42 |
| 玉米 | Zea mays | 20 |
| 金鱼草 | Antirrhinum majus | 16 |
| 陆地棉 | Gossypium hirsutum | 52 |
| 中棉 | Gossypium arboreum | 26 |
| 豌豆 | Pisum sativum | 14 |
| 香豌豆 | Lathyrus odoratus | 14 |
| 蚕豆 | Vicia faba | 12 |
| 菜豆 | phaseolus vulgaris | 22 |
| 向日葵 | Helianthusannuus | 34 |
| 烟草 | Nicotiana taldcum | 48 |
| 番茄 | Solanum lycopersicum | 24 |
| 松 | Pinus species | 24 |
| 青菜 | Brassica chinensis | 20 |
| 甘蓝 | Brassica oleracea | 18 |
| 月见草 | Oenothera biennis | 14 |
| 微生物 | ||
| 链孢霉 | Neurospora crassa | 7 |
| 青霉菌 | Penicillium species | 4 |
| 曲霉 | Aspergillus nidulans | 8 |
| 衣藻 | Chlamydomonas reinhardi | 16 |
性染色体
遗传的染色体学说的证据来自于这样的实验,一些特殊基 因的遗传行为和性染色体(Sex Chromosome)传递的关系。性染色体在高等真核生物的两种性别中是不同的。性染色体的发现为Sutton-Boveri的学说提供了一个实验证据。
因的遗传行为和性染色体(Sex Chromosome)传递的关系。性染色体在高等真核生物的两种性别中是不同的。性染色体的发现为Sutton-Boveri的学说提供了一个实验证据。
在孟德尔以前(1891年)德国的细胞学家亨金(Henking,H)曾经用半翅目的昆虫蝽做实验,发现减数分裂中雄体细胞中含11对染色体和一条不配对的单条染色体,在第一次减数分裂时,它移向一极,亨金无以为名,就称其为“X”染色体。后来在其它物种的雄体中也发现了“X”染色体。
1900年麦克朗(McClung, C.E)等就发现了决定性别的染色体。他们采用的材料多为蚱蜢和其它直翅目昆虫。1902年麦克朗发现了一种特殊的染色体,称为副染色体(accessory chromosome)。在受精时,它决定昆虫的性别。1906年威尔逊(Wilson, E.B)观察到另一种半翅目昆虫(Proteror)的雌体有6对染色体,而雄性只有5对,另外加一条不配对的染色体,威尔逊称其为X染色体,其实雌性是有一对性染色体,雄性为XO型。
在1905年斯蒂文斯(Stevens, N)发现拟步行虫属(Tenebrio molitor)中的一种甲虫雌雄个体的染色体数目是相同的,但在雄性中有一对是异源的,大小不同,其中有一条雌性中也有,但是是成对的;另一条雌性中怎么也找不到,斯蒂文斯就称之为Y染色体。在黑腹果蝇中也发现了相同的情况,果蝇共有4对染色体,在雄性中有一对是异形的染色体。在1914年塞勒(Seiler,J)证明了在雄蛾中染色体都是同形的,而在雌蛾中有一对异形染色体。他们根据异形染色体的存在和性别的相关性,发现了性染色体,已完全证实了他们的推论是完全正确的。严格地说异形染色体的存在仅是一条线索,而不是证据,不能因为存在异形染色体,就表明其为性染色体。一定要通过实验证明这条染色体上存在决定性别的主要基因,方能定论。
全面分析
2005年3月17日,在Nature杂志上发表的一篇文章宣告基本完成对人类X染色体的全面分析。对X染色体的详细测序是英国Wellcome Trust Sanger研究中心领导下世界各地多所著名学院超过250位基因组研究人员共同完成的,是人类基因组计划的一部分。
 染色体
染色体
染色体研究是临床遗传学研究的基础。测序结果表明X染色体包涵多达1100种基因。但另人吃惊的是,与之相关的疾病也有百余种,如X染色体易碎症、血友病、孤独症、肥胖肌肉萎缩病和白血病等。看来这条染色体决不容小视!
X染色体对应的另一半就是Y染色体。人类Y染色体的测序工作也已经完成,并且发现它并没有人们之前想象的那样脆弱。Y染色体上有一个“睾丸”决定基因则对性别决定至关重要。已经知道的与Y染色体有关的疾病有十几种。
异常情况
正常人的体细胞染色体数目为23对,并有一定的形态和结构。染色体在形态结构或数量上的异常被称为染色体异常,由染色体异常引起的疾病为染色体病。现已发现的染色体病有100余种,染色体病在临床上常可造成流产、先天愚型、先天性多发性畸形、以及癌肿等。染色体异常的发生率并不少见,在一般新生儿群体中就可达0.5%~0.7%,如以我院平均每年3000新生儿出生数计算,其中可能有15~20例为染色体异常者。而在早期自然流产时,约有50%~60%是由染色体异常所致。染色体异常发生的常见原因有电离辐射、化学物品接触、微生物感染和遗传等。临床上染色体检查的目的就是为了发现染色体异常和诊断由染色体异常引起的疾病。
染色体检查是用外周血在细胞生长刺激因子——植物凝集素(PHA)作用下经37℃,72小时培养,获得大量分裂细胞,然后加入秋水仙素使进行分裂的细胞停止于分裂中期前,以便染色体的观察;再经低渗膨胀细胞,减少染色体间的相互缠绕和重叠,最后用甲醇和冰醋酸将细胞固定于载玻片上,在显微镜下观察染色体的结构和数量;正常男性的染色体核型为44条常染色体加2条性染色体X和Y,检查报告中常用46,XY来表示。正常女性的常染色体与男性相同,性染色体为2条XX,常用46,XX表示。46表示染色体的总数目,大于或小于46都属于染色体的数目异常。缺失的性染色体常用O来表示。
病症示例
1. 生殖功能障碍者
在不孕症、多发性流产和畸胎等有生殖功能障碍的夫妇中至少有7%~10%是染色体异常的携带者。常见的有染色体结构变异如平衡易位和倒位以及数量异常如由于女性少一条X染色体造成的45,XO,或多一条Y染色体造成的47XXY。平衡易位和倒位由于无基因的丢失,携带者本身常并不发病,却可因其
染色体异常而导致不孕症、流产和畸胎等生殖功能障碍。性染色体数目异常除可造成不孕外,还常出现第二性征异常。
2. 第二性征异常者
常见于女性,如有原发性闭经、性发育不良,伴身材矮小、肘外翻、盾状胸和智力稍有低下,阴毛、腋毛少或缺如,后发际低,不育等,应考虑是否有X染色体异常。常见的X染色体异常有特纳氏综合征和环形X染色体。特纳氏综合征患者比正常女性少一条X染色体,其染色体核型为:45,XO。环形X染色体患者由于某种原因使X染色体两端同时出现断裂,并在断裂部位重接形成,环形染色体越小临床症状越重。早期发现这些异常并给予适当的治疗可使第二性征得到一定程度地改善,也可能获得生育能力。
3. 外生殖器两性畸形者
对于外生殖器分化模糊,如阴茎伴尿道下裂,阴蒂肥大呈阴茎样,根据生殖器外观常难以正确决定性别的患者,通过性染色体的检查有助于做出明确诊断。根据染色体检查结果和临床其它检查,两性畸形可分为真两性畸形、假两性畸形、性逆转综合征等几种不同情况。
(1)真两性畸形:内生殖器同时存在着两性的特征,即体内同时存在睾丸、输精管和卵巢、输卵管。染色体检查表现为两种类型:1、46,XX/46,XY,即一个机体内存在着两个细胞系,每种细胞的比例决定性别取向,产生的原因:X精子和Y精子同时与两个卵子受精后融合,或X精子和Y精子同时与卵细胞和刚形成、尚未排出卵外的极体分别受精所致。2、核型是46,XX,但是Y染色体的某些基因或片段易位于X染色体上,或常染色体基因突变而具有Y染色体的功能。
(2)假两性畸形:有进一步分为女假两性畸形和男假两性畸形。女假两性畸形内生殖器表现为女性,有子宫、卵巢、输卵管,染色体检查为46,XX。男性假两性畸形内生殖器表现为男性即性腺是睾丸,染色体核型是46,XY。
(3)性逆转综合征:即染色体核型与表型相反,例如核型是女性核型46,XX,但表型却似男性;或核型是男性核型46,XY,但表型却似女性。46,XX男性的主要临床表现有睾丸发育不良,隐睾,阴茎有尿道下裂,精子少或无精子,可有喉节、胡须。腋毛稀疏,群体发病率:1/2万。46,XY女性的主要临床表现有身材较高,卵巢为条索状,无子宫,盲端阴道,原发性闭经,乳房不发育。
4.先天畸形和智力低下
染色体病的特点就是多发性畸形和智力低下,常见临床特征有,头小、毛发稀而细、眼距宽、耳位低、短颈、鼻塌而短、外生殖器发育不良、腭裂、肌张低下或亢进、颠痫、通贯掌、肛门闭锁、身材矮小、发育迟缓、眼裂小、发际低、持续性新生儿黄疸及明显的青斑、眼睑下垂、心脏畸形、肾脏畸形、虹膜或视网膜缺损等。染色体检查可发现有21-三体综合征等异常。
5. 性情异常者
身材高大、性情凶猛和有攻击性行为的男性,有些可能为性染色体异常者。如XYY综合征,染色体检查表现为比正常男性多一条Y染色体,染色体核型表现为47,XYY。患者多数表型正常,即健康情况良好,常有生育能力,但子代男性中同样为47,XYY的机会大于正常人群。该病的发病率占一般男性人群的1/750。男性如出现身材修长、四肢细长、阴茎小、睾丸发不发育和精液中无精子者,有时还可以伴有智力异常,应通过染色体检查确定是否患有克氏综合症,该病患者比正常男性多一条X染色体,染色体核型表现为原`原47,XXY。其发病率在一般男性中为1/800,在男性精神发育不全者中为1%,而在男性不孕者中可高达1/10。
6. 接触过有害物质者
辐射、化学药物、病毒等可以引起染色体的断裂,如果染色体裂后原来的片段未在原来的位置上重接,将形成各种结构异常的染色体,如缺失、易位、倒位、重复、环形染色体等,这些畸变如发生在体细胞可以引起一些相应的疾病,例如肿瘤。如畸变发生在生殖细胞就发生遗传效应,殃及子代,可以引起流产、死胎、畸形儿。
7. 婚前检查
婚前检查可以发现表型正常的异常染色体携带者,如染色体平衡易位、倒位,染色体的平衡易位和倒位由于基因不丢失而表型正常,但极易引起流产、畸胎、死胎,盲目保胎会引起畸形儿的出生率增加。婚前检查还可以发现表形基本正常,但性染色体异常者,这些患者可表现为性功能障碍、无生育能力等。因此,婚前检查对优生优育有着重要的意义。
8. 白血病及其它肿瘤患者
白血病及其它肿瘤时出现的染色体异常可使血细胞的癌基因表达,使血细胞无控制的恶性生长。不同的白血病常有各自的特征性染色体异常,因此染色体检查有助于白血病的诊断和预后判定。
(1)慢性粒细胞白血病:Ph染色体是其标记染色体,由9号和22号染色体部份片段相互易位形成的。Ph染色体的出现为慢性粒细胞白血病的确诊指标,治疗过程中Ph染色体的出现或消失,还可作为疗效和愈后的参考指标。
 染色体
染色体
(3)急淋巴细胞白血病:染色体检查可发现8号和14号染色体相互易位,4号和11号染色体相互易位,9号和22号染色体相互易位形成的6条异常染色体并增加一条21号染色体。
最新进展
 染色体
染色体
中国研究人员独立完成了水稻、家蚕、鸡、吸血虫、羊等物种的全基因组测序工作。
变异情况
1、染色体变异:光学显微镜下可见染色体结构的变异或者染色体数目变异。
2、染色体结构的变异:指细胞内一个或几个染色体发生片段的缺失(染色体的某一片段消失)、增添(染色体增加了某一片段)、颠倒(染色体的某一片段颠倒了180。)或易位(染色体的某一片段移接到另一条非同源染色体上)等改变。
3、染色体数目的变异:指细胞内染色体数目增添或缺失的改变。
4、染色体组:一般的,生殖细胞中形态、大小不相同的一组染色体,就叫做一个染色体组。细胞内形态相同的染色体有几组就说明有几个染色体组。5、二倍体:凡是体细胞中含有两个染色体组的个体,就叫~。如.人果,蝇,玉米.绝大部分的动物和高等植物都是二倍体。
.6、多倍体:凡是体细胞中含有三个以上染色体组的个体,就叫~。如:马铃薯含四个染色体组叫四倍体,普通小麦含六个染色体组叫六倍体(普通小麦体细胞6n,42条染色体,一个染色体组3n,21条染色体。)。
7、一倍体:凡是体细胞中含有一个染色体组的个体,就叫一倍体。
8、单倍体:是指体细胞含有本物种配子染色体数目的个体。
9、花药离体培养法:具有不同优点的品种杂交,取F1的花药用组织培养的方法进行离体培养,形成单倍体植株,用秋水仙素使单倍体染色体加倍,选取符合要求的个体作种。
应用
1、染色体变异包括染色体结构的变异(染色体上的基因的数目和排列顺序发生改变),染色体数目变异。
2、多倍体育种:
a、成因:细胞有丝分裂过程中,在染色体已经复制后,由于外界条件的剧变,使细胞分裂停止,细胞内的染色体数目成倍增加。(当细胞有丝分裂进行到后期时破坏纺锤体,细胞就可以不经过末期而返回间期,从而使细胞内的染色体数目加倍。)
b、特点:营养物质的含量高;但发育延迟,结实率低。
c、人工诱导多倍体在育种上的应用:
常用方法---用秋水仙素处理萌发的种子或幼苗;
秋水仙素的作用---秋水仙素抑制纺锤体的形成;
实例:三倍体无籽西瓜(用秋水仙素处理二倍体西瓜幼苗得到四倍体西瓜;
用二倍体西瓜与四倍体西瓜杂交,得到三倍体的西瓜种子。三倍体西瓜联会紊乱,不能产生正常的配子。)、八倍体小黑麦。
3、单倍体育种:
形成原因:由生殖细胞不经过受精作用直接发育而成。例如,蜜蜂中的雄蜂是单倍体动物;玉米的花粉粒直接发育的植株是单倍体植物。
特点:生长发育弱,高度不孕。
单倍体在育种工作上的应用常用方法:花药离体培养法。
意义:大大缩短育种年龄。
单倍体的优点是:大大缩短育种年限,速度快,单倍体植株染色体人工加倍后,即为纯合二倍体,后代不再分离,很快成为稳定的新品种,所培育的种子为绝对纯种。
4、一般有几个染色体组就叫几倍体。如果某个体由本物种的配子不经受精直接发育而成,则不管它有多少染色体组都叫“单倍体”。
5、生物育种的方法总结如下:
①诱变育种:用物理或化学的因素处理生物,诱导基因突变,提高突变频率,从中选择培育出优良品种。实例---青霉素高产菌株的培育。
②杂交育种:利用生物杂交产生的基因重组,使两个亲本的优良性状结合在一起,培育出所需要的优良品种。实例---用高杆抗锈病的小麦和矮杆不抗锈病的小麦杂交,培育出矮杆抗锈病的新类型。
 染色体
染色体
④多倍体育种:用人工方法获得多倍体植物,再利用其变异来选育新品种的方法。(通常使用秋水仙素来处理萌发的种子或幼苗,从而获得多倍体植物。)
实例---三倍体无籽西瓜和八倍体小黑麦的培育(6n普通小麦与2n黑麦杂交得4n后代,再经秋水仙素使染色体数目加倍至8n,这就是8倍体小黑麦)。
细胞起源
染色体起源是细胞核起源的核心过程,但依然还是未解之谜。迄今为止的学说主要有:共营模型(syntrophic model)、自演化模型(autogenous model)、病毒性真核生物起源模型(viral eukaryogenesis model)、外膜假说(exomembrane hypothesis)、压缩和结构化假说(packing and structurization hypothesis),等等。
 染色体显微
染色体显微
自演化模型认为原真核(proto-eukaryotic)细胞直接自细菌演化而来,并不需要通过内共生。证据来自一类专性好氧菌——浮霉菌(Planctomycete),它们具有清晰的胞内膜结构,其中,有一种称之为Gemmata obscuriglobus的出芽菌,其染色质被双层的核膜所包裹,类似于真核生物的核的结构,而斯氏小梨形菌(Pirellula staleyi)的核被单层的细胞质内膜ICM所包裹。但是,这一模型并未进一步解释核实如何形成的。
病毒性真核生物起源模型认为,病毒感染原核生物导致了膜结合的细胞核与其他真核生物特征的产生。证据是真核生物和病毒在大分子结构上存在一定相似性,譬如,线性DNA链、mRNA的加帽,以及与蛋白质的紧密结合(病毒的外套膜类似于组蛋白)。该假说的其中一种观点认为,吞噬作用形成了早期的细胞“捕食者”,并随之演化出细胞核。
外膜假说认为,细胞核是起源自演化出第二层外细胞膜的单个早期细胞,而包裹原来细胞的内膜则转变成了核膜,并逐渐演化出精巧的核孔结构,以便于将内部(如核糖体亚基)合成的物质送出核外。
压缩与结构化假说认为,细胞核源自原核细胞基因组的大型化(包括DNA的复制错误或多倍化、侧向基因转移方式、内共生融合等)。核的成型及有丝分裂的出现主要是为了满足将巨大的DNA分子准确地分配到子代中去的需求,因此,如何将长链DNA有效地压缩(借助组蛋白)成若干染色体以及如何将多个染色体同时分离(借助纺锤体)是核演化的关键。从原核生物到真核生物,基因组的DNA总量大约增加了3.5个数量级,这与现代真核生物的DNA压缩比(packing ratio)惊人地一致。包括核膜在内的细胞内膜系统就是为了实现对复杂生化系统进行秩序化管控,或者说,秩序化是通过细胞内部的模块化得以实现的。
 平均来说,真核生物的C值比原核生物高约3.5个数量级
平均来说,真核生物的C值比原核生物高约3.5个数量级
1、本站所有文本、信息、视频文件等,仅代表本站观点或作者本人观点,请网友谨慎参考使用。
2、本站信息均为作者提供和网友推荐收集整理而来,仅供学习和研究使用。
3、对任何由于使用本站内容而引起的诉讼、纠纷,本站不承担任何责任。
4、如有侵犯你版权的,请来信(邮箱:baike52199@gmail.com)指出,核实后,本站将立即删除。













